

Ligand binding and nuclear
receptor evolution
Hector Escriva, Franck Delaunay, and Vincent Laudet*
Summary
Nuclear receptors form a superfamily of ligand-activated
transcription factors that regulate various physiological
functions, from development to homeostasis, in me-
tazoans. The superfamily contains not only receptors
for known ligands but also a large number of so-called
orphan receptors for which ligands do not exist or have
not been identified. The evolution of ligand-binding
capacity of nuclear receptors may involve either sec-
ondary loss in orphan receptors, or evolutionary acqu-
isition of ligand-binding capacity in liganded receptors.
In this review, we present arguments from phylogenetic,
functional and structural studies that support the
hypothesis that there have been several independent
gains of ligand-binding ability of nuclear receptors
during metazoan evolution. BioEssays 22:717±727,
2000. ß 2000 John Wiley & Sons, Inc.
Introduction
Nuclear hormone receptors (NRs) are one of the most
abundant classes of transcriptional regulators in metazoans,
in which they regulate functions as diverse as reproduction,
differentiation, development, metabolism, metamorphosis
and homeostasis. NRs function as ligand-activated transcrip-
tion factors, thus providing a direct link between signalling
molecules that control these processes and transcriptional
responses.
(1)
The basic concept of a NR signalling pathway
involves production of the signal (endocrine, paracrine,
autocrine or even intracrine), its transport to peripheral
organs, binding of the ligand to the receptor, and transcrip-
tional activation of the receptor.
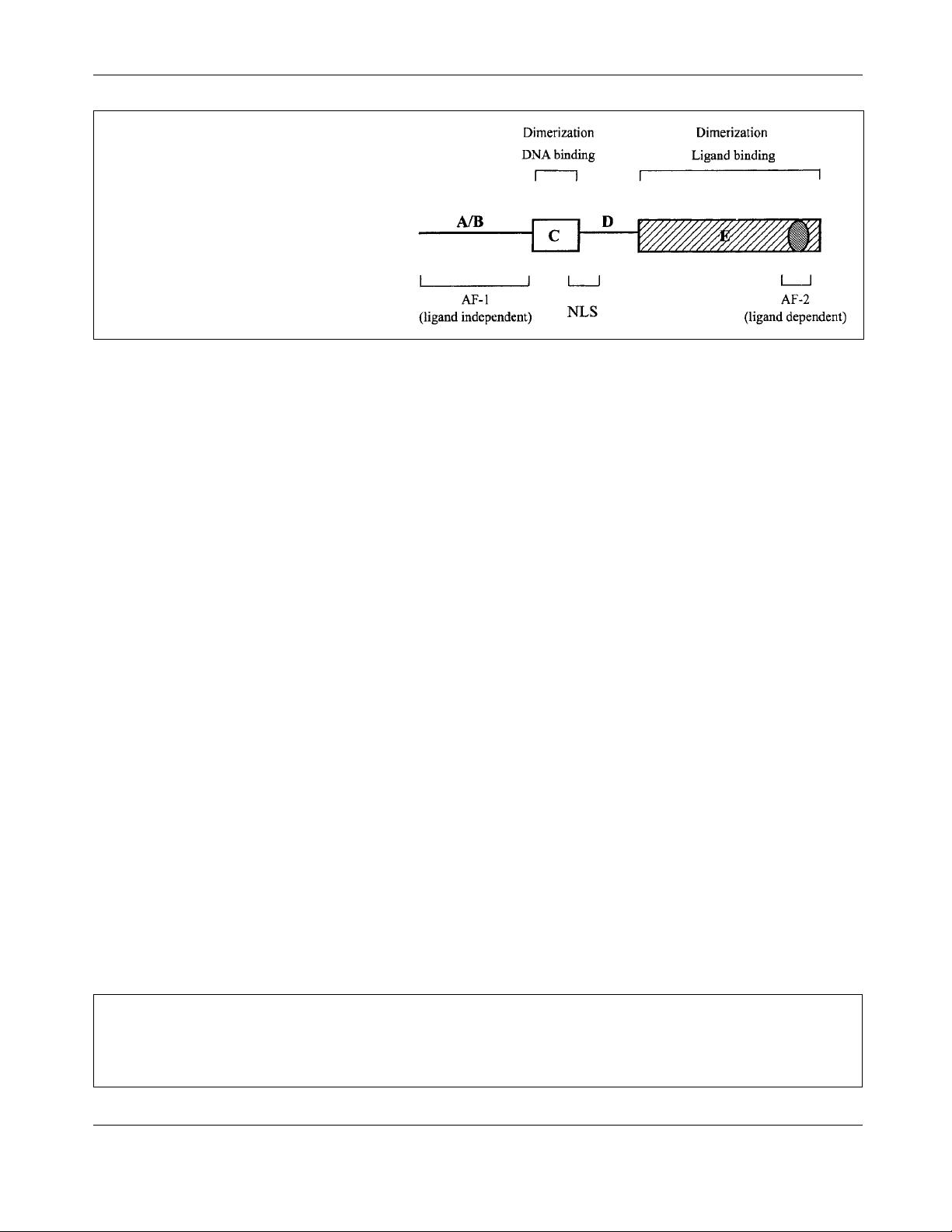
NRs share a common structural organization with a
central, well-conserved DNA-binding domain (DBD, also
termed C domain), a variable N-terminal region (A/B domain)
a non-conserved hinge (D domain) and a carboxy-terminus,
moderately conserved ligand-binding domain (LBD, E
domain) (Fig. 1).
NRs are phylogenetically related proteins clustered into a
large superfamily, including receptors for hydrophobic
molecules such as steroid hormones (estrogens, glucocorti-
coids, progesterone, mineralocorticoids, androgens, vitamin
D, ecdysone, oxysterols, bile acids etc), retinoic acids (all-
trans and 9-cis isoforms), thyroid hormones, fatty acids,
leukotrienes and protaglandins (Fig. 2). In addition, a number
of receptors, for which ligands either do not exist or still have
to be identified have been described and are referred to as
orphan receptors. NRs are promising pharmacological
targets as they bind small molecules, which can be easily
modified by drug design, and control functions associated
with major pathologies (cancer, osteoporosis, diabetes, etc).
The development of synthetic ligands for ER, RAR and
PPARg has, for instance, already led to pharmacological
applications for cancer and type 2 diabetes. Because of
these important therapeutic applications, the search of
ligands for orphan receptors and the identification of novel
signalling pathways has become a very active research
field.
(2)
The structural diversity of the ligands contrasts with the
conservation and mode of action of their receptors. This, and
the large number of orphan receptors, have prompted much
speculation on the origins of this signalling pathway. A
classical view suggests that orphan receptors have evolved
as liganded molecules, which through gene duplication
reached the present day diversity.
(3)
According to this model,
orphan receptors would have lost their ability to bind ligands
recently in evolution. We recently suggested an alternative
hypothesis, in which ligand binding was acquired during NRs
evolution.
(4)
The ancestral receptor would have been an
orphan receptor and the various liganded receptors have
gained ligand recognition independently during evolution. In
BioEssays 22:717±727, ß 2000 John Wiley & Sons, Inc. BioEssays 22.8 717
Ecole Normale Supe
Â
rieure de Lyon, Lyon, France.
Funding agencies: EMBO, Fondation pour la Recherche Me
Â
dicale,
ARC, LNCC, CNRS and MENRT.
*Correspondence to: Vincent Laudet, UMR 5665 du CNRS, Ecole
Normale Supe
Â
rieure de Lyon, 46 alle
Â
e d'Italie, 69364 Lyon Cedex 07,
France. E-mail: Vincent.Laudet@ens-lyon.fr
Abbreviations: AF2-AD, activation function 2-autonomous domain,
AR, androgen receptor, CARb, constitutive activated receptor b, DBD,
DNA-binding domain, DR, direct repeat, EcR, ecdysone receptor, ER,
estrogen receptor, ERKO, estrogen receptor knock out, ERR,
estrogen related receptor, FXR, farnesol X receptor, GCNF, germ
cell nuclear factor, GR, glucocorticoid receptor, HRE, hormone
response element, LBD, ligand-binding domain, LXR, liver X receptor,
MR, mineralocorticoid receptor, NGFIB, nerve growth factor induced
gene B, NR, nuclear receptor, PPAR, peroxisome proliferating
activated receptor, PR, progesterone receptor, PRKO, progesterone
receptor knock out, PXR, pregnane X receptor, RAR, retinoic acid
receptor, ROR, retinoic and related orphan receptor, RXR, retinoic X
receptor, SF1, steroidogenic factor 1, SRC-1, steroid receptor
coregulator 1, T3, triiodothyronine, TR, thyroid hormone receptor,
USP, ultraspiracle, VDR, vitamin D receptor.
Review articles
剩余10页未读,继续阅读
资源评论


weixin_38613173
- 粉丝: 3
- 资源: 928









最新资源
- 基于matlab的SVM二分类代码,可替数据集直接使用,算法包括数据归一化处理、c,g参数寻优,生成c,g最佳参数等高线图,交叉准确率与cg关系图及SVM预测结果图
- MATLAB制动能量回收模型simulink 轮毂电机再生制动模型 MATLAB液压电机制动模型 电动车再生制动模型 截图说明,简单易懂,一一对应 电动汽车再生制动控制策略模型,逻辑门限值控制算法
- 自动泊车APA项目开发,C代码开发泊车路径规划
- MMC型APF,MMC型statcom,MMC型储能系统,MMC-HVDC,MMC型整流器,逆变器,MMC,模块化多电平变器,MMCsimulink仿真,最近电平逼近调制,载波移相调制,排序法均衡
- a*算法与人工场势法结合的算法
- 非隔离双向DC DC变器 buck-boost变器仿真 输入侧为直流电压源,输出侧接蓄电池 模型采用电压外环电流内环的双闭环控制方式 正向运行时电压源给电池恒流恒压充电,反向运行时电池放电维持直流侧电
- 永磁同步模型电流预测控制 电流环采用预测控制双矢量改进算法 含有相关学习文献
- 行业领先的永磁同步电机pmsm 无感foc方案评估套件 套件包括电机与电路板 1、新工程采用闭环结构速度位置观测器,相比较上一代观测器; 1)低速性能进一步提升,新方案在6010电机5hz带载
- 基于Crowbar电路的双馈风力发电机DFIG低电压穿越LVRT仿真模型 Matlab Simulink仿真模型(成品) 本模型采用Crowbar保护电路(串电阻)实现低电压穿越,在电网电压跌落时投入
- 三相异步电机直接转矩DTC控制 Matlab Simulink仿真模型(成品) 传统策略DTC 1.转速环采用PI控制 2.转矩环和磁链环采用滞环控制 3.含扇区判断、磁链观测、转矩控制、开关状态选择
- 基于莱维飞行格和随机游动策略的灰狼优化算法 Matlab 源码 改进点: 1. 分段可调节衰减因子 2. 莱维飞行和随机游动策略 3. 贪婪算法寻优
- matlab simulink 在自带系统的单相MMC模型上实现了最近电平逼近调制,模型和函数是自己改的
- 松下FP系列程序 松下FP系列程序,搭配松下伺服,昆仑通态触摸屏锂电池全自动叠片贴胶机 大型程序近30000步,三个PLC,主从站通信控制 ,隔膜放卷纠偏控制,正负极真空取料叠片控制,可设定叠片
- 该模型在额定以下采用MTPA控制,速度环输出给定电流,然后代入MTPA得到dq电流,电压反馈环输出超前角进行弱磁 PI控制采用抗积分饱和,SVPWM考虑过调制情况,附带参考文献
- 自动驾驶代码-Ros移植Apollo规划方案,可编译运行,包含autoware的Lanelet2框架 帮助大家快速入门实践 完善代码,加功能等
- 一阶RC模型自适应遗忘因子递推最小二乘法+扩展卡尔曼滤波算法AFFRLS+EKF锂电池参数和SOC联合估计 遗忘因子可随时间自适应变化,不再是定值,提高估计精度 matlab程序 参考文献
资源上传下载、课程学习等过程中有任何疑问或建议,欢迎提出宝贵意见哦~我们会及时处理!
点击此处反馈


