Spiking Neural Networks and Bio-Inspired Supervised Deep Learnin
需积分: 0 116 浏览量
2023-08-02
11:28:27
上传
评论
收藏 1.04MB PDF 举报


Spiking Neural Networks and Bio-Inspired Supervised Deep Learning: A Survey
GABRIELE LAGANI
∗
, FABRIZIO FALCHI, CLAUDIO GENNARO, and GIUSEPPE AMATO, ISTI-CNR,
Italy
For a long time, biology and neuroscience elds have been a great source of inspiration for computer scientists, towards the development
of Articial Intelligence (AI) technologies. This survey aims at providing a comprehensive review of recent biologically-inspired
approaches for AI. After introducing the main principles of computation and synaptic plasticity in biological neurons, we provide a
thorough presentation of Spiking Neural Network (SNN) models, and we highlight the main challenges related to SNN training, where
traditional backprop-based optimization is not directly applicable. Therefore, we discuss recent bio-inspired training methods, which
pose themselves as alternatives to backprop, both for traditional and spiking networks. Bio-Inspired Deep Learning (BIDL) approaches
towards advancing the computational capabilities and biological plausibility of current models.
CCS Concepts: • Computing methodologies → Bio-inspired approaches; Bio-inspired approaches.
Additional Key Words and Phrases: Bio-Inspired, Hebbian, Deep Learning, Neural Networks, Spiking
ACM Reference Format:
Gabriele Lagani, Fabrizio Falchi, Claudio Gennaro, and Giuseppe Amato. 2018. Spiking Neural Networks and Bio-Inspired Supervised
Deep Learning: A Survey. 1, 1 (August 2018), 31 pages. https://doi.org/XXXXXXX.XXXXXXX
1 INTRODUCTION
Spiking Neural Networks (SNN) have recently emerged as a biologically inspired alternative to traditional Deep Learning
(DL) models, towards addressing the current limitations of Deep Neural Networks (DNNs) in terms of ecological impact
[
9
]. Indeed, biological brains exhibit extraordinary capabilities in terms of energy eciency, supporting advanced
cognitive functions while consuming only 20W [
89
]. It is believed that the key to the energy ecient computation
of biological neurons lies in the particular coding paradigm based on short pulses, or spikes [
61
]. SNN models aim at
simulating the behavior of biological neurons more realistically, compared to traditional DNNs. As a result, SNNs are
well suited for energy-ecient implementations in neuromorphic [
84
,
174
,
186
,
190
,
229
] or biological [
92
,
111
,
176
]
hardware. This makes SNNs a promising direction toward energy-ecient DL.
Unfortunately, training SNNs is not trivial, as traditional optimization based on the backpropagation algorithm
(backprop) is not directly applicable [
165
]. In fact, the biological plausibility of backprop – the workhorse of DL – is
questioned by neuroscientists [
73
,
113
,
130
,
157
,
172
]. Therefore, researchers took again inspiration from biology, in
order to nd new learning solutions as alternatives to backprop. The goal was not only to address the problem of
SNN training [
33
,
148
], but also to discover novel approaches to the learning problem [
77
,
139
,
182
], and possibly more
data ecient strategies [
69
,
90
,
105
–
107
,
110
]. In fact, another limitation of current DL solutions is the requirement of
∗
Corresp.
Authors’ address: Gabriele Lagani, gabriele.lagani@isti.cnr.it; Fabrizio Falchi, fabrizio.falchi@isti.cnr.it; Claudio Gennaro, claudio.gennaro@isti.cnr.it;
Giuseppe Amato, giuseppe.amato@isti.cnr.it, ISTI-CNR, Pisa, Italy, 56124.
Permission to make digital or hard copies of all or part of this work for personal or classroom use is granted without fee provided that copies are not
made or distributed for prot or commercial advantage and that copies bear this notice and the full citation on the rst page. Copyrights for components
of this work owned by others than ACM must be honored. Abstracting with credit is permitted. To copy otherwise, or republish, to post on servers or to
redistribute to lists, requires prior specic permission and/or a fee. Request permissions from permissions@acm.org.
© 2018 Association for Computing Machinery.
Manuscript submitted to ACM
Manuscript submitted to ACM 1
arXiv:2307.16235v1 [cs.NE] 30 Jul 2023



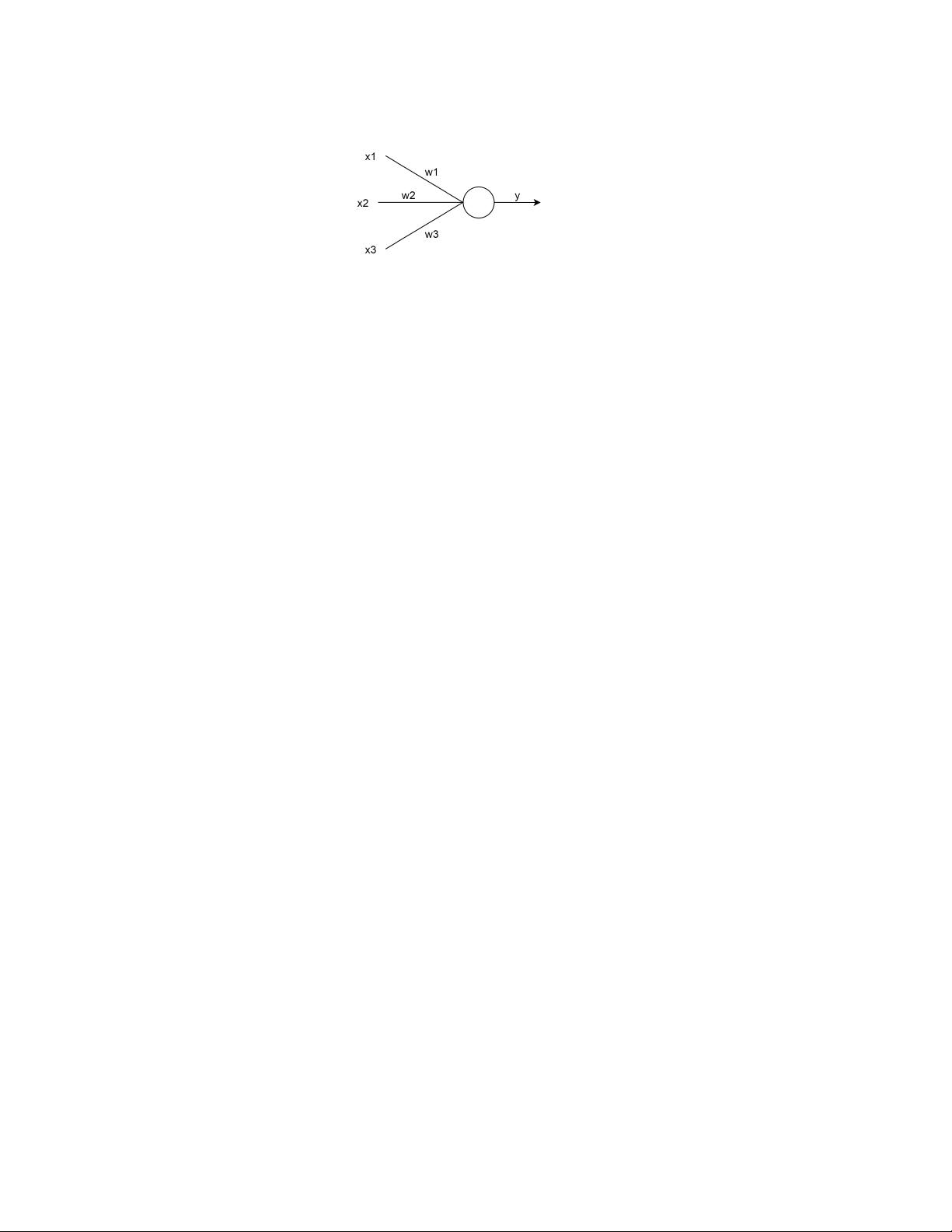
剩余30页未读,继续阅读
资源评论


学术菜鸟小晨
- 粉丝: 1w+
- 资源: 4953









最新资源
- 论文(最终)_20240430235101.pdf
- 基于python编写的Keras深度学习框架开发,利用卷积神经网络CNN,快速识别图片并进行分类
- 最全空间计量实证方法(空间杜宾模型和检验以及结果解释文档).txt
- 5uonly.apk
- 蓝桥杯Python组的历年真题
- 2023-04-06-项目笔记 - 第一百十九阶段 - 4.4.2.117全局变量的作用域-117 -2024.04.30
- 2023-04-06-项目笔记 - 第一百十九阶段 - 4.4.2.117全局变量的作用域-117 -2024.04.30
- 前端开发技术实验报告:内含4四实验&实验报告
- Highlight Plus v20.0.1
- 林周瑜-论文.docx
资源上传下载、课程学习等过程中有任何疑问或建议,欢迎提出宝贵意见哦~我们会及时处理!
点击此处反馈


