

1. 引言
实现移动机器人的高精度定位逐渐成为机器人导航重要的研究方向之一
[1,2]
,也是实现
机器人智能化决策和控制的基础。通过传感器获取信息和基于概率的同步定位与地图构建
(Simultaneous Localization And Mapping, SLAM)算法是目前机器人自主导航的主要研究方
向。科学家通过对生物环境认知能力的研究发现,哺乳类动物具有对环境的自主认知能
力。O’Keefe 等人
[3]
在对鼠类海马体椎体神经细胞的放电率进行研究时发现,一些椎体神经
细胞的放电活动具有明显的位置选择性,即当大鼠到达某一特定位置时,对应的神经细胞
产生复杂的尖峰电位,而其余细胞处于抑制状态。O’Keefe 等人将具有这种特性的细胞称
为位置细胞(Place Cell, PC)。Hafting 等人
[4]
发现放电形式为六边形的网格细胞,多个网格细
胞的关联响应可以实现空间导航。在之后的研究中,O’Keefe 等人和挪威的 Moser 等人
[5-7]
根据位置细胞和网格细胞具有的“定位”属性,提出了大脑里的“导航系统”概念。文献[8]对
海马体位置细胞的功能和特性作出了更加详细的说明。更多的研究发现,通过对位置细胞
放电率的计算,结合路标获取、位置编码和图像信息能完成机器人的空间位置估计
[9,10]
。空
军工程大学的李伟龙等人
[11]
基于位置细胞的放电机理,提出了位置细胞表征空间位置的两
种方法,借助环境路标的提取实现了对于机器人空间位置的高精度表征。周牧等人
[12]
提出
一种基于 Mann-Whitney 秩和检验的无线局域网室内映射与定位方法,该方法可根据实际
定位精度需求对目标区域进行分段处理,在无需运动传感器辅助和构建位置指纹数据库的
条件下,实现更高的映射与定位精度,但存在相同聚类接收信号强度(Received Signal
Strength, RSS)样本冗余等问题导致聚类(即信号空间中的骨干节点)映射到物理空间中的邻
接性较差。文献[13]提出的基于 RSS 非齐性分布特征的半监督学习室内定位指纹库构建算
法,实现运用少量标记数据对大量未标记数据的位置标定,但仍存在高维接收信号强度
(RSS)数据分布不均匀的影响,缺乏对环境局部几何特征的提取性能。Aronov 等人
[14]
在大
鼠奖励实验中发现,当大鼠到达“奖励空间”时,位于海马体的位置细胞活动速率达到最
大,将这些活动速率变大的位置细胞的响应位置记录下来,得到一条覆盖大鼠活动区域的
路径,即大鼠的运动路径。Milford 等人
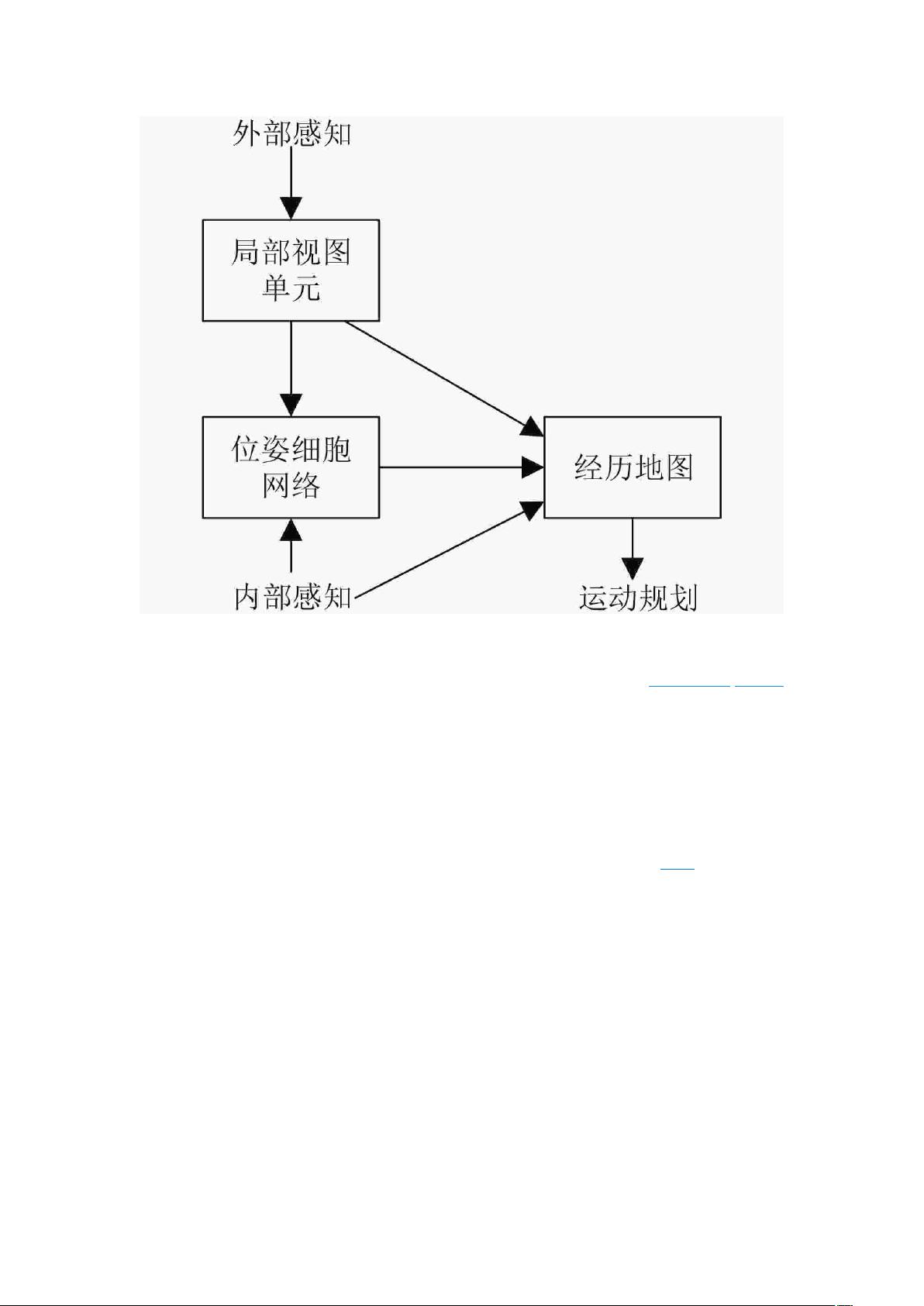
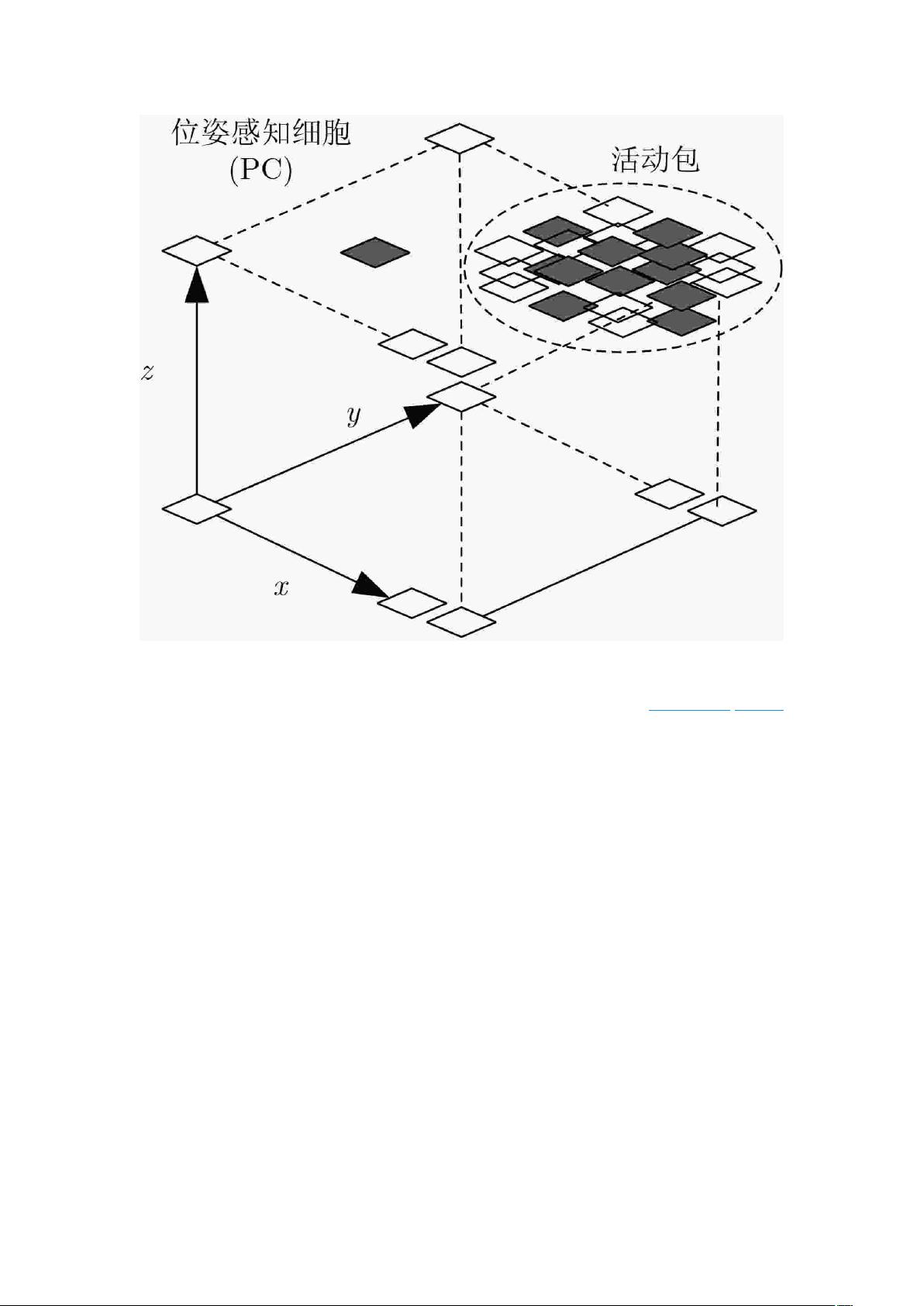
[15-17]
提出了一种通过关联视觉场景细胞和位姿细胞
建立经历地图的仿生 SLAM 算法,这一算法能有效地进行闭环检测,更快实现空间地图构
建。但也存在定位精度低和处理数据速度慢的缺点。
1982 年芬兰科学家 Kohonen
[18,19]
基于侧抑制现象提出一种自组织图(Self-Organizing
Map, SOM),并引入“赢者通吃”的竞争规则。这种自组织方法通过学习能够自主绘制出拓
扑地图,但必须定义其初始网络结构,无法满足系统实时性要求。Montazeri 等人
[20]
提出一
种将增长自组织特征图应用于强化学习的算法,通过两个增长的自组织映射实现状态空间
的最佳表征。于乃功等人
[21]
根据自组织可增长映射图(Growing Self-Organizing Map, GSOM)

剩余20页未读,继续阅读
资源评论


罗伯特之技术屋
- 粉丝: 4525
- 资源: 1万+

下载权益

C知道特权

VIP文章

课程特权
开通VIP









最新资源
- Java毕业设计-springboot-vue-工作量统计系统(源码+sql脚本+29页零基础部署图文详解+29页论文+环境工具+教程+视频+模板).zip
- Java毕业设计-springboot-vue-高校疫情防控web系统(源码+sql脚本+29页零基础部署图文详解+31页论文+环境工具+教程+视频+模板).zip
- 基于模型的六轴机器人阻抗控制算法演示与仿真参数设置指导(matlab simscape仿真机器人模型自定义切换,跟踪轨迹展示及算法学习),基于模型的六轴机器人阻抗力控制算法(matlab simsca
- Java毕业设计-springboot-vue-工资信息管理系统(源码+sql脚本+29页零基础部署图文详解+25页论文+环境工具+教程+视频+模板).zip
- 基于PaddlePaddle的眼疾识别项目(包含训练集、测试集、模型权重)
- Java毕业设计-springboot-vue-航班进出港管理系统(源码+sql脚本+29页零基础部署图文详解+29页论文+环境工具+教程+视频+模板).zip
- 基于ABAQUS、Opensees和Perform3d的结构易损性评估与IDA曲线绘制:matlab函数调用与地震波选波调幅实战指南,ABAQUS 、Opensees、Perform3d IDA曲线
- Java毕业设计-springboot-vue-果蔬作物疾病防治系统(源码+sql脚本+29页零基础部署图文详解+30页论文+环境工具+教程+视频+模板).zip
- Java毕业设计-springboot-vue-滑雪场管理系统(源码+sql脚本+29页零基础部署图文详解+27页论文+环境工具+教程+视频+模板).zip
- Java毕业设计-springboot-vue-家教管理系统(源码+sql脚本+29页零基础部署图文详解+32页论文+环境工具+教程+视频+模板).zip
- Java毕业设计-springboot-vue-驾校预约学习系统(源码+sql脚本+29页零基础部署图文详解+34页论文+环境工具+教程+视频+模板).zip
- 数据采集系统:下位机与上位机协同工作,高性能模数转换与灵活配置滤波功能,数据采集系统下位机与上位机代码 下位机采用开发板来完成 AD9226模数转芯片,最大65MHz采样,12bit量化 双通
- Java毕业设计-springboot-vue-集团门户网站(源码+sql脚本+29页零基础部署图文详解+33页论文+环境工具+教程+视频+模板).zip
- 蓝桥杯Java历年真题及其解析.docx
- Java毕业设计-springboot-vue-教师薪酬管理系统(源码+sql脚本+29页零基础部署图文详解+32页论文+环境工具+教程+视频+模板).zip
- Java毕业设计-springboot-vue-教学辅助系统(源码+sql脚本+29页零基础部署图文详解+31页论文+环境工具+教程+视频+模板).zip
资源上传下载、课程学习等过程中有任何疑问或建议,欢迎提出宝贵意见哦~我们会及时处理!
点击此处反馈


